Gynaikothrips uzeli (Zimmermann, 1900)
Phlaeothripinae, Phlaeothripidae, Tubulifera, Thysanoptera
Figures
Fig. 1: 8-segmented antenna, segments III and IV with simple sense cones, terminal segments VI-VIII
Fig. 2: Head dorsal with ocellar triangle
Fig. 3: Head ventral with mouth cone
Fig. 4: Pronotum
Fig. 5: Meso- and metanotum
Fig. 6: Pelta
Fig. 7: Tergites IV and V with wing-retaining setae
Fig. 8: Tergite X (= tube)
Introduction and recognition
Gynaikothrips uzeli is a major pest of Ficus species, particularly Ficus benjamina. Both sexes fully winged. Body dark brown; tarsi and apices of tibiae yellow; also antennal segments II-V yellow, VI shaded at apex, VII shaded in distal half and VIII light brown (Fig. 1); fore wings pale with margins shaded on distal half. Antennae 8-segmented; segment III with 1 sense cone, IV with 3 sense cones. Head longer than wide, slightly constricted behind eyes (Fig. 2); first ocellus raised on a prominence over the antennal bases; postocular setae with apices bluntly pointed, arising well behind compound eyes (Fig. 2), scarcely extending to posterior margin of eye; maxillary stylets retracted almost to postocular setae, about one third of head width apart. Pronotum with strongly developed transverse lines of sculpture forming conspicuous swirls; with major setae variable, anteromarginals minute, anteroangulars commonly well developed, posteroangulars at least 0.7 times as long as the epimeral setae, and always longer than the discal setae (Fig. 4); epimeral sutures variably complete. Prosternal basantra not present; mesopraesternum broadly boat-shaped; metathoracic sternopleural sutures not developed. Metanotum strongly sculptured with longitudinal reticulations (Fig. 5). Fore tarsus with small or minute tooth. Fore wing parallel sided; with about 15 duplicated cilia. Pelta broadly triangular, median reticulations with internal markings (Fig. 6); tergites II-VII with 2 pairs of sigmoid wing-retaining setae (Fig. 7); tergite IX setae S1 acute and about 0.8 as long as tube; tube longer than head (Fig. 8).
Male smaller than female; no fore tarsal tooth; tergite IX setae S2 short and stout; sternite VIII without glandular area.
Taxonomic identity
Species
Gynaikothrips uzeli (Zimmermann, 1900)
Taxonomic history
Gynaikothrips garitacambroneroi Retana-Salazar, 2006
Phloeothrips longitubus Bagnall, 1909
Mesothrips uzeli Zimmermann, 1900
Common name
Cuban laurel thrips
Weeping fig thrips
Present taxonomic position
Family: Phlaeothripidae Uzel, 1895
Subfamily: Phlaeothripinae Uzel, 1928
Genus: Gynaikothrips Zimmermann, 1900
Genus description
The genus Gynaikothrips Zimmermann, 1900
This genus contains 40 species, which feed on leaves and produce leaf-galls or leaf-rolls. Most of these species are found in the Oriental region on tropical or subtropical plant material, but 2 related pest species Gynaikothrips ficorum and Gynaikothrips uzeli are distributed worldwide due to horticultural trade with ornamental Ficus spp. (Mound & Kibby 1998). These are large thrips species with dark bodies, sometimes containing internal pigmentation, and 8-segmented antennae. Members of this genus have 1 sense cone on antennal segment III and 3 on segment IV, and due to these characters is thought to be related to the genus Liothrips.
Species description
Typical key character states of Gynaikothrips uzeli
Coloration
Body color: mainly brown to dark brown
Antennae
Number of antennal segments: 8
Form of sensorium on antennal segments III and IV: emergent and simple on segments III and IV
Segment III - number of sense cones: 1
Segment IV - number of sense cones: 3
Head
Maxillary stylet position: about one third of head width apart
Postocular setae: about half to nine tenths as long as compound eye
Maxillary bridge: absent
Cheeks: without seta-bearing tubercle
Prothorax
Prosternal basantra: absent
Epimeral suture: interrupted
Pronotal posteroangular and epimeral setae: posteroangular seta at least 0.7 times as long as the epimeral setae (always longer than the discal setae)
Wings
Fore- and hind wings: present, more than half as long as abdomen
Fore wing veins: absent
Fore wing shape: mainly parallel sided or margins run continuously towards each other
Fore- and hind wing surface: not covered with microtrichia, smooth
Fore wing surface: not reticulate
Fringe cilia arising: not from sockets
Fore wing duplicate cilia: present
Fore wings: uniformly pale or weakly shaded
Fore wing extreme apex color: pale
Legs
Fore tarsus: without a tooth or hook
Abdomen
Tergites III to V: with two pairs of sigmoid curved wing-retaining seta
Abdominal segment 10: complete tube in both sexes
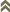
Similar or related species
Gynaikothrips uzeli is very similar in structural details to Gynaikothrips ficorum. The only reported difference between these two species is the length of the pronotal posteroangular pair of setae. In Gynaikothrips uzeli, these setae are at least 0.7 times as long as the epimeral setae, and always longer than the discal setae. In Gynaikothrips ficorum, these setae are less than 0.5 times as long as the epimerals and usually no longer than the discal setae. A more practical, but less accurate, way to distinguish Gynaikothrips uzeli from Gynaikothrips ficorum is by host plant association. Mound et al. (1995) suggest that Gynaikothrips uzeli is the primary gall maker (i.e., leaf folder) on Ficus benjamina, whereas Gynaikothrips ficorum is the primary gall maker (i.e., leaf roller) on Ficus microcarpa. Mound, Wang & Okajima (1995) suggested that Gynaikothrips ficorum is probably a form of Gynaikothrips uzeli that has been widely distributed by the horticultural trade. Further studies, especially on the ITS-RFLP-patterns of these two species should clarify the species state, but first results show very similar patterns of DNA-fragments using both "sub-species".
Compared to other members of Phlaeothripidae, species of Gynaikothrips have parallel sided fore wings (other species with fore wings distinctly constricted medially, only Karnyothrips flavipes with weakly constricted fore wings). Almost all species have a mainly brown body color, uniformly fore wings and tergites II-VII each with 2 pairs of wing-retaining setae (only Aleurodothrips fasciapennis with a bicolored body, fore wings with transverse alternating bands of dark and light, and tergites II-VII each with 1 pair of wing-retaining setae), and no seta-bearing tubercle on cheeks (only Hoplandrothrips marshalli has cheeks with at least 1 seta-bearing tubercle in basal third or 3 pairs laterally). Species of Gynaikothrips are similar to Karnyothrips flavipes and species of Haplothrips in having their maxillary stylet apart by about one third of head width apart, and the length of postocular setae is about half to nine tenths as long as eye (in Aleurodothrips fasciapennis the maxillary stylet position is more than half of head width apart, and postocular setae are shorter than distance of the setal base from the eye; in Hoplandrothrips marshalli the maxillary stylets are about one fifth of head width apart, and postocular setae are as long as dorsal length of eye or a little longer). Species of Gynaikothrips as well as Hoplandrothrips marshalli and Aleurodothrips fasciapennis have neither a maxillar bridge nor a prosternal basantra (Karnyothrips flavipes and species of Haplothrips with maxillary bridge and prosternal basantra). Like Hoplandrothrips marshalli, Karnyothrips flavipes and Haplothrips (Haplothrips) gowdeyi, species of Gynaikothrips have duplicated cilia on posterior margin of fore wing (Aleurothrips fasciapennis and Haplothrips (Trybomiella) clarisetis without duplicated cilia on posterior margin of fore wing).
Biology
Life history
Eggs are laid in large numbers inside the surface of the curled leaf. Development from egg to fertile females takes about 30 days. All stages of development can be found at any time within the pocket galls.
Host plants
Ficus benjamina is the only plant on which it is known to complete its life cycle. Other recorded hosts include F. obtusa, F. pilosa, F. microcarpa.
Vector capacity
None identified, but possible mechanical distribution of phytopathogenic fungi and bacteria.
Damage and symptoms
According to Mound et al. (1995), Gynaikothrips ficorum is the primary gall maker on Ficus microcarpa while Gynaikothrips uzeli is the primary gall maker on F. benjamina. Adult feeding induces the young foliage of the host to fold along the midvein, forming a gall. This folded gall is distinctly different from the rolled galls induced by Gynaikothrips ficorum on its primary host, Ficus microcarpa. Foliage infested with Gynaikothrips uzeli becomes galled and appear distorted and gnarled. Infestations cause premature leaf drop and stunted growth.
Detection and control strategies
Biological control: Dozier (1926) reported two species of Anthocoridae (minute pirate bugs), Macrotracheliella laevis and Cardiastethus rugicollis, to be predacious on Cuban laurel thrips in Puerto Rico and should be effective against Gynaikothrips uzeli. The minute pirate bug Montandoniola moraguesi and the eulophid wasp Thripastichus gentile, both are predatory insects specialized on gall inducing thrips (Dobbs & Boyd 2006; La Salle 1994).
Cultural control: Gynaikothrips uzeli only attacks the tender new foliage on small plants. It should be possible to prune out the new growth and eliminate the thrips population. Consequently there is no suitable foliage for feeding and oviposition and the infestation should die out before new growth emerges. Use of a resistant species of Ficus is also recommended.
Additional notes
-
Biogeography
Southeast Asia (including Taiwan, China, India), Indonesia, North America, Hawaii, Trinidad, Costa Rica, Kenya.
African countries where Gynaikothrips uzeli has been reported

Occurence of Gynaikothrips uzeli in East Africa

Please click here for survey sites of all observed thrips species of Kenya, Tanzania and Uganda.
Click here for locations of Gynaikothrips uzeli in parts of East Africa.
Bibliography
Ananthakrishnan TN (1978). Thrips galls and gall thrips. Technical Monograph, Zoological Survey of India. 1: 1-62
Bagnall RS (1909). On some new and little known exotic Thysanoptera. Transactions of the Natural History Society of Northumberland. 3: 524-540
Dobbs TT & Boyd Jr. DW (2006). Status and distribution of Montandoniola moraguesi (Hemiptera: Anthocoridae) in the continental United States. Florida Entomologist. 89 (1): 41-46
Dozier HL (1926). Notes on Puerto Rican Thysanoptera. Journal of the Department of Agriculture of Puerto Rico. 10 (3-4): 279-281
Held DW & Boyd Jr. DW (2008). Evaluation of sticky traps and insecticides to prevent gall induction by Gynaikothrips uzeli Zimmerman (Thysanoptera: Phlaeothripidae) on Ficus benjamina. Pest Management Science. 64 (2): 133-140
Held DW & Boyd Jr. DW (2008). New records of Gynaikothrips uzeli (Zimmerman) (Thysanoptera: Phlaeothripidae) on Ficus benjamina in Texas and O'ahu, Hawaii, U.S.A. Pan-Pacific Entomologist. 84 (2): 77-80
Held DW, Boyd D, Lockley T & Edwards GB (2005). Gynaikothrips uzeli (Thysanoptera: Phlaeothripidae) in the Southeastern United States: Distribution and review of biology. Florida Entomologist. 88 (4): 538-540
Karny H, van Leeuwen-Reijnvaan W & van Leeuwen-Reijnvaan J (1916). Beiträge zur Kenntnis der Gallen von Java. Zweite Mitteilung über die javanischen Thysanopterocecidien und deren Bewohner. Zeitschrift für Wissenschaftliche Insektenbiologie. 12: 84-94
La Salle J (1994). North American genera of Tetrastichinae (Hymenoptera: Eulophidae). Journal of Natural History. 28 (1): 109-236
Lewis T (1973). Thrips: their biology, ecology and economic importance. Academic Press Inc., London Ltd., 349 pp
Lewis T (1997). Thrips as crop pests. CAB International, Wallingford, 740 pp
Moritz G (2006). Thripse. Pflanzensaftsaugende Insekten, Bd. 1, (1. Auflage). Westarp Wissenschaften, Hohenwarsleben, 384 pp. ISBN 13: 978 3 89432 8917
Moritz G, Morris DC & Mound LA (2001). ThripsID - Pest thrips of the world. ACIAR and CSIRO Publishing Collingwood, Victoria, Australia, CDROM ISBN 1 86320 296 X
Mound LA (1968). A review of R. S. Bagnalľs Thysanoptera collections. Bulletin of the British Museum (Natural History), Entomology. Supplement 11: 1-181
Mound LA & Kibby G (1998). Thysanoptera: An identification guide, (2nd edition). CAB International, Wallingford and New York, 70 pp
Mound LA & Marullo R (1996). The thrips of Central and South America: An introduction (Insecta: Thysanoptera). Memoirs on Entomology, International, Vol. 6. Associated Publishers, Gainsville, 487 pp
Mound LA, Wang C-L & Okajima S (1995). Observations in Taiwan on the identity of the Cuban laurel thrips (Thysanoptera, Phlaeothripidae). Journal of the New York Entomological Society. 103 (2): 185-190
Priesner H (1939). Zur Kenntnis der Gattung Gynaikothrips Zimm. (Thysanoptera). Mitteilungen der Münchner Entomologischen Gesellschaft. 29: 475-487
Retana-Salazar AP (2006). Variación morfológica del complejo Gynaikothrips uzeli-ficorum (Phlaeothripidae: Tubulifera). Métodos Ecologia Sistemética. 1 (1): 1-9
Wheeler C, Held DW & Boyd Jr. DW (2006). Morphological differences in between two gall-inducing species, Gynaikothrips uzeli and Gynaikothrips ficorum. Proceedings of the Southern Nursery Association Research Conference (Section 3 - Entomology). 51: 153-155
Wolfenbarger DO (1946). Cuban laurel thrips control on Ficus benjamini. Florida Entomologist. 28 (4): 82-83
Zimmermann A (1900). über einige javanische Thysanoptera. Bulletin de ľInstitut Botanique de Buitenzorg. 7: 6-19
----
Web links
Mound´s Thysanoptera pages
Thysanoptera Checklist
ICIPE Thrips survey sites
UNI Halle & Thrips sites
Thrips of California









